Type
Recombinant protein
Description
Total 192 AA. Mw: 21.2 kDa (calculated). UniProtKB acc.no. Q9JJN1. N-terminal His-tag, 10 extra AA.
Amino Acid Sequence
MKHHHHHHASAYPIPDSSPLLQFGGQVRQRYLYTDDDQDTEAHLEIREDGTVVGAAHRSPESLLELKALKPGVIQILGVKASRFLCQQPDGALYGSPHFDPEACSFRELLLEDGYNVYQSEAHGLPLRLPQKDSPNQDATSWGPVRFLPMPGLLHEPQDQAGFLPPEPPDVGSSDPLSMVEPLQGRSPSYAS
Source
E. coli
Purity
˃ 90 % by SDS-PAGE
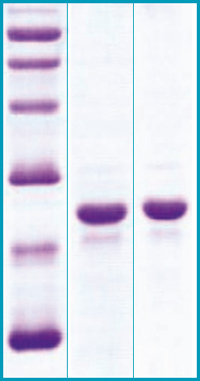
SDS-PAGE Gel
12% SDS-PAGE separation of Mouse FGF21
1. M.W. marker – 14, 21, 31, 45, 66, 97 kDa
2. reduced and heated sample, 5μg/lane
3. non-reduced and non-heated sample, 5μg/lane
Endotoxin
< 1.0 EU/µg
Formulation
Filtered (0,4 μm) and lyophilized in 0.5 mg/mL in 20mM TRIS, 50mM NaCl, pH 7.5
Reconstitution
Add deionized water to prepare a working stock solution of approximately 0.5 mg/mL and let the lyophilized pellet dissolve completely. Add DTT (0.2 mM ) and NaCl (0.1–0.15 M) before freezing to prevent potential aggregation.
Applications
Western blotting, ELISA
Shipping
At ambient temperature. Upon receipt, store the product at the temperature recommended below.
Storage/Expiration
Store the lyophilized protein at -80 °C. Lyophilized protein remains stable until the expiry date when stored at -80 °C. Aliquot reconstituted protein to avoid repeated freezing/thawing cycles and store at -80 °C for long term storage. Reconstituted protein can be stored at 4 °C for a week.
Quality Control Test
BCA to determine quantity of the protein.
SDS PAGE to determine purity of the protein.
Endotoxin level determination.
Note
This product is intended for research use only.
Research topic
Diabetology - Other Relevant Products, Energy metabolism and body weight regulation, Animal studies
Summary
The FGFs are a family of more than 20 small (~17–26 kDa) secreted peptides. The initial characterization of these proteins focused on their ability to stimulate fibroblast proliferation. This mitogenic activity was mediated through FGF receptors (FGFRs) 1, 2, or 3. A fourth closely related tyrosine kinase receptor (FGFR4) was able to bind the FGFs but did not lead to a mitogenic response.
FGFs modulate cellular activity via at least 5 distinct subfamilies of high-affinity FGF receptors (FGFRs): FGFR-1, –2, –3, and –4, all with intrinsic tyrosine kinase activity and, except for FGFR-4, multiple splice isoforms, and FGFR-5, which lacks an intracellular kinase domain. There is growing evidence that FGFRs can be important for regulation of glucose and lipid homeostasis. The overexpression of a dominant negative form of FGFR-1 in β cells leads to diabetes in mice, which thus implies that proper FGF signaling is required for normal β cell function and glycemia maintenance. FGFR-2 appears to be a key molecule during pancreatic development. Moreover, FGFR-4 has been implicated in cholesterol metabolism and bile acid synthesis.
FGF-19, has been shown to cause resistance to diet-induced obesity and insulin desensitization and to improve insulin, glucose, and lipid profiles in diabetic rodents. Since these effects, at least in part, are mediated through the observed changes in metabolic rates, FGF-19 can be considered as a regulator of energy expenditure.
FGF-21 is preferentially expressed in liver, but an exact knowledge of FGF-21 bioactivity and its mode of action have been lacking to date. FGF-21 is a potent activator of glucose uptake on adipocytes, protects animals from diet-induced obesity when overexpressed in transgenic mice, and lowers blood glucose and triglyceride levels when therapeutically administered to diabetic rodents.