Type
Recombinant protein
Amino Acid Sequence
ETTTQGPGVLLPLPKGAATGWMAGIPGHPGHNGAPGRDGRDGTPGEKGEKGDPGLIGPKGDIGETGVPGAEGPRGFPGIQGRKGEPGEGAYVYRSAFSVGLETYVTIPNMPIRFTKIFYNQQNHYDGSTGKFHCNIPGLYYFAYHITVYMKDVKVSLFKKDKAMLFTYDQYQENNVDQASGSVLLHLEVGDQVWLQVYGEGERNGLYADNDNDSTFTGFLLYHDTNDYKDDDDK
Source
HEK293
Purity
˃ 90 % by SDS-PAGE
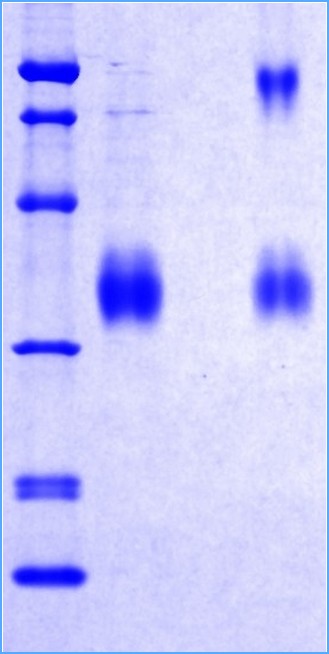
SDS-PAGE Gel
12% SDS-PAGE separation of Human Adiponectin, Trimeric form
1. M.W. marker – 14, 21, 31, 45, 66, 97 kDa
2. reduced and heated sample, 5μg/lane
3. non-reduced and non-heated sample, 5μg/lane
Endotoxin
< 1.0 EU/µg
Formulation
Filtered (0.4 μm) and lyophilized from 0.5 mg/ml solution in phosphate buffered saline pH 7.5
Reconstitution
Add deionized water to prepare a working stock solution of approximately 0.5 mg/mL and let the lyophilized pellet dissolve completely.
Applications
Western blotting, ELISA
Shipping
At ambient temperature. Upon receipt, store the product at the temperature recommended below.
Storage/Expiration
Store the lyophilized protein at -80 °C. Lyophilized protein remains stable until the expiry date when stored at -80 °C. Aliquot reconstituted protein to avoid repeated freezing/thawing cycles and store at -80 °C for long term storage. Reconstituted protein can be stored at 4 °C for a week.
Quality Control Test
BCA to determine quantity of the protein.
SDS PAGE to determine purity of the protein.
Endotoxin level determination.
Note
This product is intended for research use only.
Research topic
Chronic renal failure, Coronary artery disease, Diabetology - Other Relevant Products, Energy metabolism and body weight regulation
Summary
Adiponectin, also referred to as Acrp30, AdipoQ and GBP-28, is a recently discovered 244 aminoacid protein, the product of the apM1 gene, which is physiologically active and specifically and highly expressed in adipose cells. The protein belongs to the soluble defence collagen superfamily; it has a collagen-like domain structurally homologous with collagen VIII and X and complement factor C1q-like globular domain. Adiponectin forms homotrimers, which are the building blocks for higher order complexes found circulating in serum. Together, these complexes make up approximately 0.01% of total serum protein. Adiponectin receptors AdipoR1 and AdipoR2 have been recently cloned; AdipoR1 is abundantly expressed in skeletal muscle, whereas AdipoR2 is predominantly expressed in the liver. Paradoxically, adipose tissue-expressed adiponectin levels are inversely related to the degree of adiposity. Adiponectin concentrations correlate negatively with glucose, insulin, triglyceride concentrations, liver fat content and body mass index and positively with high-density lipoprotein-cholesterol levels, hepatic insulin sensitivity and insulin-stimulated glucose disposal. Adiponectin has been shown to increase insulin sensitivity and decrease plasma glucose by increasing tissue fat oxidation. Of particular interest is that low adiponectin serum levels predict type 2 diabetes independent of other risk factors. Adiponectin also inhibits the inflammatory processes of atherosclerosis suppressing the expression of adhesion and cytokine molecules in vascular endothelial cells and macrophages, respectively. This adipokine plays a role as a scaffold of newly formed collagen in myocardial remodelling after ischaemic injury and also stimulates angiogenesis by promoting cross-talk between AMP-activated protein kinase and Akt signalling in endothelial cells. Low serum adiponectin levels are found in patients with coronary artery disease. Moreover, high circulating levels of adiponectin are associated with decreased risk of myocardial infarction, independent of other factors. Altogether, adiponectin has the potential to become a clinically relevant parameter to be measured routinely in subjects at risk for type 2 diabetes, atherosclerosis and the metabolic syndrome.